Lipopolysaccharide tolerance attenuates inflammatory responses by increasing heme oxygenase-1 and tristetraprolin expression in Raw264.7 macrophages
Abstract
Inflammation is associated with the progression of a variety of diseases. Lipopolysaccharides (LPS) tolerance is recognized to reduce proinflammatory responses. Heme oxygenase1 (HO1) and tristetraprolin (TTP) are induced by LPS tolerance and mediate the anti-inflammatory effects. However, it was not clear whether two molecules are linked in LPS tolerance. In this study, we sought to determine whether HO1 associates with TTP to mediate the anti-inflammatory effects of LPS tolerance. LPS treatment significantly increased mRNA and protein level of HO1 and TTP in a time dependant fashion while LPS significantly decreased mRNA and protein level of TNFα in Raw264.7 macrophages. LPS tolerance inhibited TNFα mRNA and protein while HO1 and TTP level was still increased. In HO1 deficient macrophages, LPS tolerance failed to attenuate TNFα mRNA expression but TTP level was still decreased. Our results suggest that HO1 and TTP are functionally linked in mediating anti-inflammatory effects of LPS tolerance. This novel LPS tolerance-HO1-TTP signaling pathway provides new possibilities for the treatment of inflammatory diseases.
INTRODUCTION
Inflammation is associated with the progression of a variety of diseases such as cancer, atherosclerosis, asthma, psoriasis, and so on. Lipopolysaccharides (LPS), secreted by gram-negative bacteria activates macrophages. Activated macrophages mediate excessive production of proinflammatory cytokines including tumor necrosis factor α (TNFα), which leads to serious systemic disorders. Pretreatment of LPS is involved with reduction of sensitivity to subsequent challenge of LPS which is termed LPS tolerance. LPS tolerance was observed in vivo as well as in vitro with a reduced production of inflammatory cytokines in response to a secondary LPS stimulation. However, the precise mechanisms involved in LPS tolerance-induced alteration of the proinflammatory responses remains unclear.
The inflammatory response modulated by post-transcriptional control [1, 2] is dependent on AU-rich element (ARE)–mediated mechanisms [3-5]. The ARE-binding protein, tristetraprolin (TTP) binds and destabilizes pro-inflammatory mRNAs including TNFα [6-9]. The importance of TTP in inhibiting inflammatory responses has been implicated in TTP-deficient mice that develop severe inflammatory arthritis, autoimmune dysfunction, and myeloid hyperplasia [10]. These mice produce increasing amount of the proinflammatory cytokine TNFα by macrophages and suffer from slow growth, underweight and the development of polyarticular arthritis. In mouse macrophages, increase in TTP expression and activity was associated with decrease expression of TNFα mRNA [11]. In addition, increase in TTP mRNA expression in response to LPS stimulation was observed in dendritic cells.
It was reported that LPS treatment induces the expression of TTP [12, 13]. LPS-induced TTP seems to exert an inhibitory function against LPS-induced lung inflammation, because TTP deficiency aggravates LPS-induced lung inflammation.
Hemeoxygenase 1 (HO1) is a stress-inducible protein and the anti-inflammatory effects of HO1 may have therapeutic potential in inflammatory conditions such as arthritis [14]and inflammatory bowel disease [15]. HO1 and TTP are functionally linked in mediating the anti-inflammatory effects of nicotine; HO1 is necessary for the induction of TTP. Increased HO1 expression was observed in the lung during LPS tolerance and cross tolerance [16]. In addition, overexpression of hepatic HO1 has been observed during LPS tolerance [17]. Currently, there are no reports regarding the effects of HO1 on the regulation of TTP expression on LPS tolerance under pro-inflammatory conditions.
Therefore, in our study, we focused on detailed mechanisms under laying the LPS tolerance effects on HO1- TTP expression in macrophages.
MATERIALS AND METHODS
Reagents and antibodies
Anti-β-actin, anti-TTP, anti-HO1 and horseradish peroxidase conjugated secondary (anti-mouse IgG and anti-goat IgG) antibodies and small interfering RNA (siRNAs) against mouse HO1, and control siRNA were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). LPS and protease inhibitor cocktail sets were purchased from Sigma Aldrich (St. Louis, MO, USA). Dulbecco’s Modified Egale Medium (DMEM), fetal bovine serum (FBS), penicillin–streptomycin and sodium pyruvate were purchased from Invitrogen (Grand Island, NY, USA). All other chemicals were purchased from Sigma-Aldrich.
Cell culture
Mouse Raw264.7 macrophage cells were cultured in DMEM supplemented with 10% FBS and 1% penicillin-streptomycin. Cells were seeded in 6-well plates at a density of 5×105/ml in DMEM medium supplemented with 10% FBS. Supernatants were collected.
Transfection
Cells (5×105/ml) were cultured in 6-well plate for 18h and transfected with HO1 siRNA (100 nM) using Lipofectamine 2000 (Invitrogen, CA, USA). Cells were pretreated with LPS 100 ng/ml for 24 h in the presence or absence of 1 μg/ml LPS for 6 h. The expression levels of HO1, TTP, or TNFα mRNA were analyzed by q-RT-PCR.
Western blot assays
Cell lysates were prepared using RIPA buffer containing protease inhibitors and phosphatase inhibitors and total protein concentration of the lysates was measured using a BCA Protein Assay kit (Pierce Biotechnology Inc., Rockford, IL, USA). Proteins were resolved by SDS-PAGE, transferred onto polyvinylidenedifluoride (PVDF) membrane, and probed with appropriate dilutions of the following antibodies: anti-TTP, anti-HO1, and anti-β-actin. Immunoreactivity was detected using the ECL detection system (GE healthcare Bio-Sciences Corp, NJ). Films were exposed at multiple time points to ensure that the images were not saturated.
Reverse transcription-polymerase chain reaction (RT-PCR)
Total RNA was extracted using TRIzol reagent (Invitrogen) according to manufacturer’s instructions. DNase I-treated total RNA (2 μg) was reversed transcribed using M-MLV reverse transcriptase (Promega Corporation, WI, USA) and oligo-dT (Promega Corporation, WI, USA). Semi-quantitative RT-PCR was performed using Taq polymerase (Solgent, Daejeon, Korea) and PCR primer pairs as follows: GAPDH: 5′-aggccggtgctgagtatgtc-3′, 5′-tgcctgcttcaccttct-3′; HO-1: 5′-tcccagacaccgctcctccag-3′, 5′-ggatttggggctggtttc-3′; TTP: 5′-ctctgccatctacgagagcc-3′, 5′-gatggagtccgagtttatgttcc-3′; TNFα, 5′-agcccacgtcgtagcaaaccaccaa-3′, 5′-acacccattcccttcacagagcaat-3′. The gene amplification reaction conditions were as follows: denaturation at 94ºC for 0.5 min; annealing at 58-62ºC (based on the melting temperature of each respective primer) for 0.5 min; extension at 72ºC for 1 min: PCR cycles were determined according to a kinetic profile. GAPDH was used as an internal loading control.
Enzyme linked immunosorbent assay (ELISA)
TNFa in the cell supernatants were analyzed using Mouse TNF-alpha Quantikine ELISA Kit (R&D Systems).
Statistical analysis
Statistical differences between groups were evaluated by one-way ANOVA or student’s t-test. A p value of <0.05 was considered statistically significant.
RESULTS
LPS increases HO1 and TTP expression in macrophages
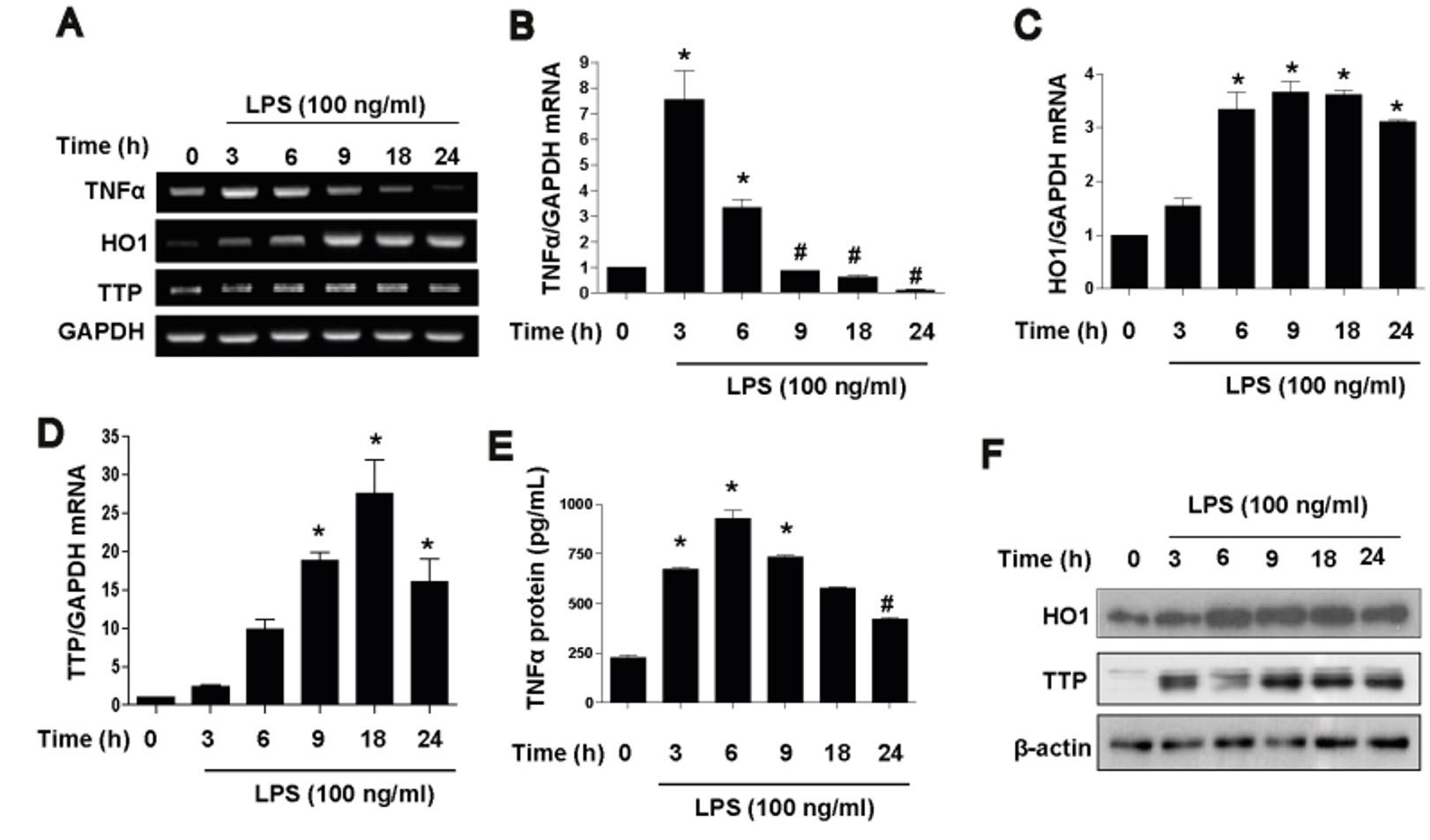
HO1 is a well known anti-inflammatory protein and TTP is known to destabilization of TNFα mRNA ARE sites. To prepare LPS-induced endotoxin tolerance in vitro model we treated Raw264.7 macrophages with LPS in a time dependent manner. LPS treatment significantly increased mRNA and protein level of HO1 (Figure 1A, C and F) and TTP (Figure 1A, D and F) in a time dependant fashion. Conversely, LPS significantly decreased mRNA and protein level of TNFα in a time dependant manner (Figure 1A, B and E). These events were more significant at the time of 18 or 24 h.
LPS tolerance inhibits inflammatory responses via HO1-TTP expression in macrophages
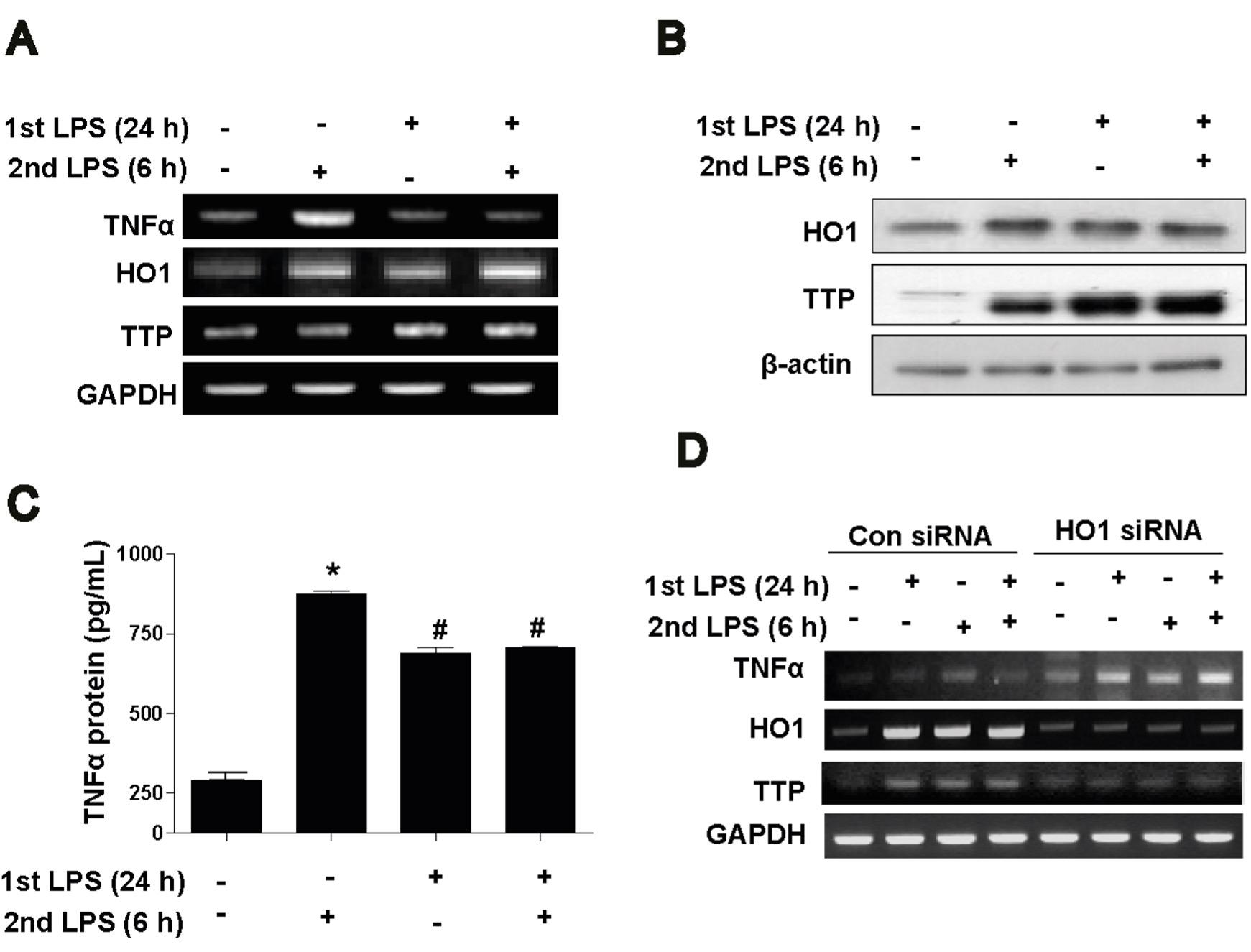
To determine whether LPS tolerance has any effect on TTP expression as well as inflammatory responses. We pretreated Raw264.7 macrophages with LPS for 24 h (1st hit as LPS tolerant model) and again stimulated with LPS for 6 h (2nd hit). Results showed that LPS tolerance prevented effect of 2nd challenge of LPS as measured by TNFα mRNA and protein (Figure 2 A and C) while HO1 and TTP level was still increased (Figure 2 A and B). To further confirm the effect of TTP in LPS tolerance, we used HO1 siRNA. In con siRNA, LPS tolerance limited effects of 2nd hit of LPS as shown by TNFα mRNA expression (Figure 2D) but in HO1 siRNA, LPS tolerance failed to suppress TNFα mRNA expression (Figure 2D) while HO1 siRNA effectively suppressed HO1 and TTP expression which indicating the link between HO1 and TTP as described previously. Evidences and our data indicate that LPS tolerance in macrophages is mediated by HO1-TTP axis.
DISCUSSION
Endotoxin tolerance induced by LPS preconditioning is recognized to reduce proinflammatory responses. In our study, we reported that LPS tolerance attenuates inflammatory responses through increasing HO1-TTP system.
Molecular mechanisms for LPS tolerance have long been investigated [18]. LPS-signaling pathways have been shown to be affected by several proteins. In addition to HO1, TTP is induced by LPS treatment [13]. HO1 and TTP provide protection against inflammatory diseases and LPS tolerance was found to increase their expression [13, 17]. Thus, it is hypothesized that LPS tolerance increases the expression of HO1 and TTP and thus, suppresses inflammatory responses. TTP suppresses inflammation by destabilizing the mRNAs of pro-inflammatory cytokines. LPS-induced TTP seems to exert an inhibitory function against LPS-induced lung inflammation, because TTP deficiency aggravates LPS-induced lung inflammation. HO1 mediates its anti-inflammatory effects through TTP [19]. Despite evidence for the roles of HO1 and TTP in LPS tolerance-mediated anti-inflammatory effects, the functional links between HO1 and TTP in endtoxin tolerance state is unclear. In this report, we demonstrate that increased expression of HO1 and TTP functions in the endtoxin tolerance -mediated anti-inflammatory signaling pathway. Our data provide evidence for the requirement of HO1 in LPS tolerance-induced upregulation of TTP in macrophages. In addition, silencing of HO1 blocks the effect of LPS tolerance on the induction of TTP and inhibits LPS-induced TNFα production both in vitro. Collectively, our data suggest that HO1 and TTP mediate the anti-inflammatory function of LPS tolerance in the HO1-TTP signaling pathway.
In summary, these results suggest that HO1 and TTP are functionally linked in mediating the anti-inflammatory function of LPS tolerance. LPS tolerance enhances the expression of HO1-TTP levels which in turn inhibits LPS-induced TNFa production. This study identified a novel LPS tolerance-HO1-TTP signaling pathway responsible for the inhibition of LPS-driven inflammation and potentially provides the rationale for novel therapeutic approaches for the management of inflammatory diseases.
ACKNOWLEDGEMENT
The author thanks to Dr. Md Jamal Uddin for his kind help during experiments and manuscript preparation.
CONFLICT OF INTERESTS
The author declares no conflict of interest.
References
- [1]Stoecklin G, Anderson P. Posttranscriptional mechanisms regulating the inflammatory response. AdvImmunol. 2006;89:1-37.
- [2]Hollams EM, Giles KM, Thomson AM, Leedman PJ. MRNA stability and the control of gene expression: implications for human disease. Neurochem Res. 2002;27:957-80.
- [3]Clark A. Post-transcriptional regulation of pro-inflammatory gene expression. Arthritis Res. 2000;2:172-4.
- [4]Kracht M, Saklatvala J. Transcriptional and post-transcriptional control of gene expression in inflammation. Cytokine. 2002;20:91-106.
- [5]Khabar KS. The AU-rich transcriptome: more than interferons and cytokines, and its role in disease. J Interferon Cytokine Res. 2005;25:1-10.
- [6]Carballo E, Lai WS, Blackshear PJ. Feedback inhibition of macrophage tumor necrosis factor-alpha production by tristetraprolin. Science. 1998;281:1001-5.
- [7]Phillips K, Kedersha N, Shen L, Blackshear PJ, Anderson P. Arthritis suppressor genes TIA-1 and TTP dampen the expression of tumor necrosis factor alpha, cyclooxygenase 2, and inflammatory arthritis. Proc Natl Acad Sci U S A. 2004;101:2011-6.
- [8]Park D, Lee EK, Jang EJ, Jeong HO, Kim BC, Ha YM, et al. Identification of the dichotomous role of age-related LCK in calorie restriction revealed by integrative analysis of cDNA microarray and interactome. Age (Dordr).
- [9]Qian X, Ning H, Zhang J, Hoft DF, Stumpo DJ, Blackshear PJ, et al. Posttranscriptional regulation of IL-23 expression by IFN-gamma through tristetraprolin. J Immunol.186:6454-64.
- [10]Taylor GA, Carballo E, Lee DM, Lai WS, Thompson MJ, Patel DD, et al. A pathogenetic role for TNF alpha in the syndrome of cachexia, arthritis, and autoimmunity resulting from tristetraprolin (TTP) deficiency. Immunity. 1996;4:445-54.
- [11]Anderson P. Post-transcriptional control of cytokine production. Nat Immunol. 2008;9:353-9.
- [12]Brooks SA, Connolly JE, Rigby WF. The role of mRNA turnover in the regulation of tristetraprolin expression: evidence for an extracellular signal-regulated kinase-specific, AU-rich element-dependent, autoregulatory pathway. J Immunol. 2004;172:7263-71.
- [13]Chen YL, Jiang YW, Su YL, Lee SC, Chang MS, Chang CJ. Transcriptional regulation of tristetraprolin by NF-kappaB signaling in LPS-stimulated macrophages. Mol Biol Rep. 2013;40:2867-77.
- [14]Devesa I, Ferrandiz ML, Terencio MC, Joosten LA, van den Berg WB, Alcaraz MJ. Influence of heme oxygenase 1 modulation on the progression of murine collagen-induced arthritis. Arthritis Rheum. 2005;52:3230-8.
- [15]Hegazi RA, Rao KN, Mayle A, Sepulveda AR, Otterbein LE, Plevy SE. Carbon monoxide ameliorates chronic murine colitis through a heme oxygenase 1-dependent pathway. J Exp Med. 2005;202:1703-13.
- [16]Koch A, Boehm O, Zacharowski PA, Loer SA, Weimann J, Rensing H, et al. Inducible nitric oxide synthase and heme oxygenase-1 in the lung during lipopolysaccharide tolerance and cross tolerance. Crit Care Med. 2007;35:2775-84.
- [17]Soriano RN, Ravanelli MI, Batalhao ME, Carnio EC, Branco LG. Glucocorticoids downregulate systemic nitric oxide synthesis and counteract overexpression of hepatic heme oxygenase-1 during endotoxin tolerance. Can J Physiol Pharmacol. 2013;91:861-5.
- [18]Ziegler-Heitbrock HW. Molecular mechanism in tolerance to lipopolysaccharide. J Inflamm. 1995;45:13-26.
- [19]Jamal Uddin M, Joe Y, Zheng M, Blackshear PJ, Ryter SW, Park JW, et al. A functional link between heme oxygenase-1 and tristetraprolin in the anti-inflammatory effects of nicotine. Free Radic Biol Med. 2013;65:1331-9.