Resveratrol attenuates inflammation through tristetraprolin expression in human hepatocytes
Abstract
Resveratrol (Resv), a polyphenolic compound has the anti-inflammatory and antioxidant effects. Tristetraprolin (TTP) destabilizes mRNAs of pro-inflammatory cytokines. However, it was not clear how Resv exerts anti-inflammatory effects on hepatocytes. In this study, we used human hepatoma cell line (HepG2 cells) and analyzed samples using qRT-PCR. Resv increases TTP expression in hepatocytes. Resv attenuates LPS-induced inflammation through TTP expression in hepatocytes. In TTP deficient hepatocytes, LPS failed to attenuate TNFα, iNOS, and IL-6 mRNA expression whereas TTP level was still decreased. Our results suggest that Resv may attenuate inflammation in hepatocytes through TTP induction at least in part. This novel Resv-TTP signaling pathway provides new possibilities for the treatment of inflammatory diseases in liver.
INTRODUCTION
Resveratrol (Resv; 3,5,4′-trihydroxy-trans-stilbene), a polyphenolic antioxidant compound derived from grape skin is present in red wine and certain medicinal plants. Recently, it is found to possess hepatoprotective effect [1]. It provides protection against various infections and stress [2]. It also various kind of pharmacological effects against heart disorders, lipoprotein oxidation, free radical, apoptosis, as well as platelet aggregation [3]. Further, it can defend against EtOH-induced lipid peroxidation and protects liver in rat [4].
Inflammation is associated with the progression of different kinds of diseases. Gram-negative bacteria activate macrophages by secreting lipopolysaccharides (LPS). Activated macrophages mediate excessive production of proinflammatory cytokines including tumor necrosis factor α (TNFα), which leads to serious systemic disorders. The antioxidant and anti-inflammatory effects of Resv has been established [5]. Resv inhibits the increased levels of TNFα and interleukin (IL)-6 in diabetic kidneys [6].
The inflammatory response modulated by post-transcriptional control [7] is dependent on AU-rich element (ARE)–mediated mechanisms [8]. The ARE-binding protein, tristetraprolin (TTP) binds and destabilizes pro-inflammatory mRNAs including TNFα [9]. TTP-deficient mice show severe inflammatory arthritis, autoimmune dysfunction, and myeloid hyperplasia [10], indicating the importance role of TTP in inhibiting inflammatory responses. These mice produce increasing amount of the proinflammatory cytokine TNFα by macrophages leading to polyarticular arthritis.
It is also reported that LPS treatment induces the expression of TTP [11]. LPS-induced TTP seems to exert an inhibitory function against LPS-induced lung inflammation, because TTP deficiency aggravates LPS-induced lung inflammation. Resv stimulates HO-1 expression and this activation is related to the anti-inflammatory and antioxidant effects [12,13]. HO1 and TTP are functionally linked in mediating the anti-inflammatory effects; HO1 is necessary for the induction of TTP [14,15]. These observations suggest that Resv may have protective function in inflammation through TTP induction in hepatocytes.
Therefore, in our study, we used Resv to check the effect of TTP on LPS-induced inflammation in HepG2 cells.
MATERIALS AND METHODS
Reagents and antibodies
Small interfering RNA (siRNAs) against human TTP, and control siRNA were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). LPS was purchased from Sigma Aldrich (St. Louis, MO, USA). Dulbecco’s Modified Egale Medium (DMEM), fetal bovine serum (FBS), penicillin–streptomycin and sodium pyruvate were purchased from Invitrogen (Grand Island, NY, USA). All other chemicals were purchased from Sigma-Aldrich.
Cell culture
The human hepatocellular carcinoma cell line, HepG2, was purchased from ATCC (Manassas, VA, USA) and maintained with DMEM supplemented with 10% fetal bovine serum (FBS), 1% penicillin/streptomycin. Cells were seeded in 6-well plates at a density of 5×105/ml.
Transfection
HepG2 cells (5×105/ml) were cultured in 6-well plate for 18h and transfected with TTP siRNA (100 nM) using Lipofectamine 2000 (Invitrogen, CA, USA). Cells were pretreated with or without Resv and stimulated in the presence or absence of LPS. The expression levels of TTP, TNFα, iNOS, and IL-6 mRNA were analyzed by q-RT-PCR.
Reverse transcription-polymerase chain reaction (RT-PCR)
Total RNA was extracted using TRIzol reagent (Invitrogen) according to manufacturer’s instructions. DNase I-treated total RNA (2 μg) was reversed transcribed using M-MLV reverse transcriptase (Promega Corporation, WI, USA) and oligo-dT (Promega Corporation, WI, USA). Real-time PCR was performed using SYBR Green PCR Master Mix (QIAGEN) on an ABI 7500 Fast Real-Time PCR System (Applied Biosystems). Semi-quantitative RT-PCR was performed using Taq polymerase (Solgent, Daejeon, Korea). PCR primer pairs for human were as follows: TTP: 5-cgctacaagactgagctat-3, 5-gag-gtagaacttgtgacaga-3; TNF-α: 5’-tcacaagcacctcccaaagc-3’, 5’-agcaagctgtgccactttcc-3’; iNOS: 5’-gcagaatgtgaccatcatgg-3’, 5’-acaaccttggtgttgaaggc-3’; IL-6: 5’-aagccagagctgtgcagatgagta-3’, 5’-cttggtcaccgacgtcctgt-3’; GAPDH: 5’-acatcaagaaggtggtgaag-3’, 5’-ctgttgctgtagccaaattc-3’. PCR cycles were determined according to a kinetic profile. GAPDH was used as an internal loading control.
Statistical analysis
Statistical differences between groups were evaluated by one-way ANOVA or student’s t-test. A p value of <0.05 was considered statistically significant.
RESULTS
Resveratrol increases TTP expression in hepatocytes
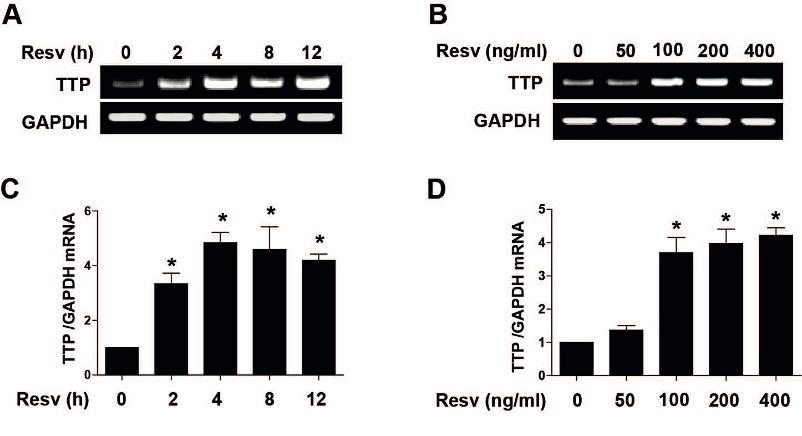
To explore the effects of Resv on TTP expression, time and dose dependent treatment of Resv was employed in HepG2 cells. Resv significantly increased mRNA of TTP at different time points (0, 2, 4, 8, and 12 h) by using RT-PCR (Figure 1A) and qRT-PCR (Figure 1C). Based on TTP mRNA expression patterns, we used 4 h treatment time for out subsequent experiments. Similarly, Resv significantly increased mRNA of TTP at different doses (0, 100, 200, and 400 ng/ml) by using RT-PCR (Figure 1B) and qRT-PCR (Figure 1D). These data suggest that Resv increases TTP expression in human hepatocytes.
Resveratrol attenuates LPS-induced inflammation through TTP expression in hepatocytes
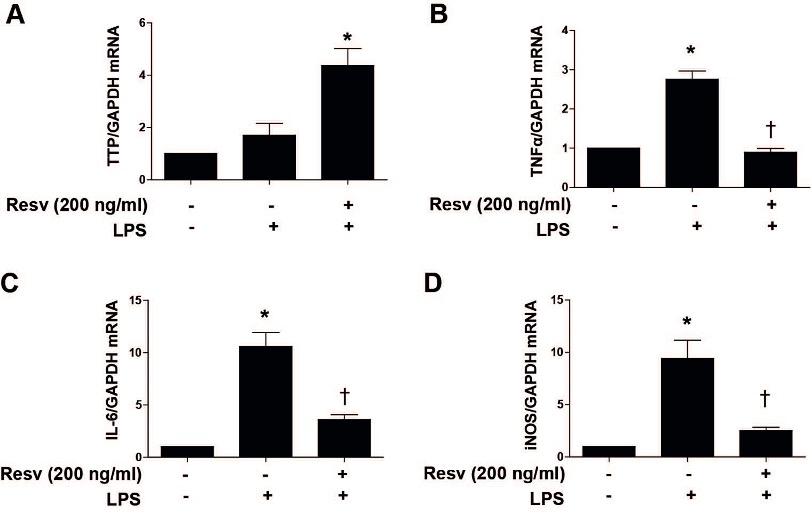
To determine whether Resv has any effect on LPS-induced pro-inflammatory cytokine expression as well as TTP expression. HepG2 cells were pretreated with Resv for 4 h and again stimulated with LPS for 6 h. Results showed that Resv significantly increased TTP expression (Figure 2A) as well as decreased pro-inflammatory cytokines such as TNFα, IL-6, and iNOS (Figure 2B-D) in LPS-stimulated cells.
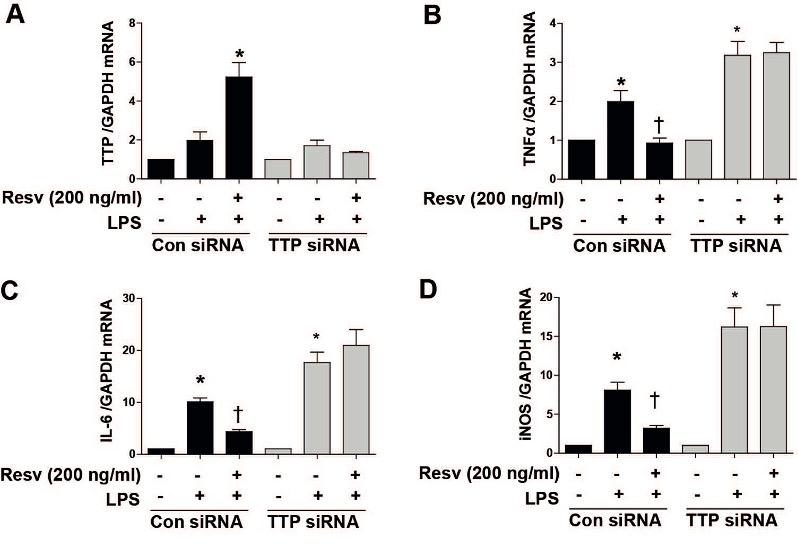
To confirm the involvements of TTP on inhibition of pro-inflammatory cytokines, we used TTP siRNA. Under con siRNA, treatment of Resv significantly increased TTP mRNA expression while under TTP siRNA, Resv failed to increase the mRNA level of TTP in LPS-stimulated hepatocytes (Figure 3A), suggesting the effectiveness of transfection. Further, we checked the role of TTP on pro-inflammatoy cytokines by using TTP siRNA. We found that Resv significantly inhibited the pro-inflammatory cytokines such as TNFα, IL-6, and iNOS under con siRNA, where these were not affected by TTP siRNA (Figure 3B-D). Data suggest that Resv-induced TTP may have anti-inflammatory effects in human hepatocytes.
DISCUSSION
Resv has well-known antioxidant and anti-inflammatory properties and exerts numerous pharmacological effects including hepatoprotection. The present study provides the direct evidence that TTP activation by Resv represents a novel therapeutic mechanism by which TTP prevents LPS-induced inflammation in hepatocytes.
Resv significantly increased mRNA of TTP at different time points and doses in HepG2 cells. These data suggest that Resv increases TTP expression in human hepatocytes. In addition, Resv attenuated LPS-induced inflammation through TTP expression in hepatocytes. Consistently, TTP activation by Resv inhibits the proliferation and metastasis of colorectal cancer cells [16] and induces glioma cell apoptosis [17].
LPS-signaling pathways have been shown to be affected by several proteins. TTP suppresses inflammation by destabilizing the mRNAs of pro-inflammatory cytokines [7]. LPS-induced TTP seems to exert an inhibitory function against LPS-induced lung inflammation, because TTP deficiency aggravates LPS-induced lung inflammation. On the other hand, HO1 is a stress-inducible protein and the anti-inflammatory effects of HO1 may have therapeutic potential in inflammatory conditions such as arthritis [18] and inflammatory bowel disease [19]. However, HO1 and TTP are functionally linked in mediating the anti-inflammatory effects [14]. Recently, our group report that the effects of HO1 on the regulation of TTP expression on LPS tolerance under pro-inflammatory conditions. In our study, Resv significantly inhibited the pro-inflammatory cytokines such as TNFα, IL-6, and iNOS under con siRNA, where these were not affected by TTP siRNA, suggest that Resv-induced TTP may have anti-inflammatory effects in human hepatocytes.
However, the major limiting factor in preclinical dissolution testing for drug and health food applications, the water solubility and adsorption of Resv is poor [20]. Therefore, red wine is not recommended for patients with alcohol-related disorders due to poor dissolution property and adsorption of Resv [21].
In summary, these results suggest that Resv may attenuate inflammation in hepatocytes through TTP induction at least in part. This study identified a novel Resv-TTP signaling pathway responsible for the inhibition of LPS-driven inflammation and potentially provides the rationale for novel therapeutic approaches for the management of inflammatory liver disorders.
ACKNOWLEDGEMENT
This work is supported by Korea Research Fellowship Program through National Research Foundation (NRF) funded by Korean Ministry of Education, Science, and Technology (2015H1D3A1062189).
AUTHOR CONTRIBUTIONS
AM designed and performed the experiments. AM and MJU wrote the manuscript. AM and AI analyzed the data.
CONFLICT OF INTEREST
The authors declare that no conflicts of interest exist.
References
- [1]Liu S, Hou W, Yao P, Zhang B, Sun S, Nüssler AK, et al. Quercetin protects against ethanol-induced oxidative damage in rat primary hepatocytes. Toxicol Vitr 2010; 24(2):516-522. doi:10.1016/j.tiv.2009.03.006.
- [2]Gonthier B, Allibe N, Cottet-Rousselle C, Lamarche F, Nuiry L, Barret L. Specific conditions for resveratrol neuroprotection against ethanol-induced toxicity. J Toxicol 2012; 2012:973134. doi:10.1155/2012/973134.
- [3]Shakibaei M, Harikumar KB, Aggarwal BB. Review: Resveratrol addiction: To die or not to die. Mol Nutr Food Res 2009. doi:10.1002/mnfr.200800148.
- [4]Kasdallah-Grissa A, Mornagui B, Aouani E, Hammami M, Gharbi N, Kamoun A, et al. Protective effect of resveratrol on ethanol-induced lipid peroxidation in rats. Alcohol Alcohol 2006; 41(3):236-239. doi:10.1093/alcalc/agh256.
- [5]Frémont L. Biological effects of resveratrol. Antioxid Redox Signal 2001. doi:10.1089/152308601317203567.
- [6]Kumar A, Sharma SS. NF-κB inhibitory action of resveratrol: A probable mechanism of neuroprotection in experimental diabetic neuropathy. Biochem Biophys Res Commun 2010. doi:10.1016/j.bbrc.2010.03.014.
- [7]Stoecklin G, Anderson P. Posttranscriptional Mechanisms Regulating the Inflammatory Response. Adv Immunol 2006. doi:10.1016/S0065-2776(05)89001-7.
- [8]Carpenter S, Ricci EP, Mercier BC, Moore MJ, Fitzgerald KA. Post-transcriptional regulation of gene expression in innate immunity. Nat Rev Immunol 2014. doi:10.1038/nri3682.
- [9]Carballo E, Lai WS, Blackshear PJ. Feedback inhibition of macrophage tumor necrosis factor-alpha production by tristetraprolin. Science 1998. doi:10.1126/science.281.5379.1001.
- [10]Taylor GA, Carballo E, Lee DM, Lai WS, Thompson MJ, Patel DD, et al. A pathogenetic role for TNFα in the syndrome of cachexia, arthritis, and autoimmunity resulting from tristetraprolin (TTP) deficiency. Immunity 1996. doi:10.1016/S1074-7613(00)80411-2.
- [11]Mahtani KR, Brook M, Dean JL, Sully G, Saklatvala J, Clark AR. Mitogen-activated protein kinase p38 controls the expression and posttranslational modification of tristetraprolin, a regulator of tumor necrosis factor alpha mRNA stability. Mol Cell Biol 2001. doi:10.1128/MCB.21.9.6461.
- [12]Zhuang H, Kim YS, Koehler RC, Doré S. Potential mechanism by which resveratrol, a red wine constituent, protects neurons. Ann. N. Y. Acad. Sci., 2003. doi:10.1111/j.1749-6632.2003.tb07534.x.
- [13]Juan S-H, Cheng T-H, Lin H-C, Chu Y-L, Lee W-S. Mechanism of concentration-dependent induction of heme oxygenase-1 by resveratrol in human aortic smooth muscle cells. Biochem Pharmacol 2005. doi:10.1016/j.bcp.2004.09.015.
- [14]Jamal Uddin M, Joe Y, Zheng M, Blackshear PJ, Ryter SW, Park JW, et al. A functional link between heme oxygenase-1 and tristetraprolin in the anti-inflammatory effects of nicotine. Free Radic Biol Med 2013. doi:10.1016/j.freeradbiomed.2013.09.027.
- [15]Akhi Moni. Lipopolysaccharide tolerance attenuates inflammatory responses by increasing hemeoxygenase 1 and tristetraprolin expression in Raw264.7 macrophages. J Adv Biotechnol Exp Ther 2018;1:07-10. doi:10.5455/jabet.2018.d5.
- [16]Lee SR, Jin H, Kim WT, Kim WJ, Kim SZ, Leem SH KS. Tristetraprolin activation by resveratrol inhibits the proliferation and metastasis of colorectal cancer cells. Int J Oncol 2018. doi:10.3892/ijo.2018.4453.
- [17]Ryu J, Yoon NA, Seong H, Jeong JY, Kang S, Park N, et al. Resveratrol Induces Glioma Cell Apoptosis through Activation of Tristetraprolin. Mol Cells 2015. doi:10.14348/molcells.2015.0197.
- [18]Devesa I, Ferrándiz ML, Terencio MC, Joosten LAB, Van Den Berg WB, Alcaraz MJ. Influence of heme oxygenase 1 modulation on the progression of murine collagen-induced arthritis. Arthritis Rheum 2005. doi:10.1002/art.21356.
- [19]Hegazi RAF, Rao KN, Mayle A, Sepulveda AR, Otterbein LE, Plevy SE. Carbon monoxide ameliorates chronic murine colitis through a heme oxygenase 1–dependent pathway. J Exp Med 2005. doi:10.1084/jem.20051047.
- [20]Rodrigo R, Miranda A, Vergara L. Modulation of endogenous antioxidant system by wine polyphenols in human disease. Clin Chim Acta 2011. doi:10.1016/j.cca.2010.11.034.
- [21]Lee CW, Yen FL, Huang HW, Wu TH, Ko HH, Tzeng WS, et al. Resveratrol nanoparticle system improves dissolution properties and enhances the hepatoprotective effect of resveratrol through antioxidant and anti-inflammatory pathways. J. Agric. Food Chem., 2012. doi:10.1021/jf2050137.